Trap Plants
Description
This section is from the book "The Psychic Life Of Insects", by E. L. Bouvier. Also available from Amazon: The Psychic Life of Insects.
Trap Plants
We have for a, long time given the name trap plants to a certain number of Asele-piads and Zingïberace whose flowers are so made that they often hold in fatal captivity the insects that visit them. Kùnckel d 'Her-culais (1909) has studied the mechanism of this capture in an Argentine Ascle-piad, Araujia sericofera, and in a magnificent Zingiber of the genus HedycJi-ium. In the first the beak is wedged at the summit of the narrow angular space which leave between them the hardened expansions of the two contiguous stamens. In the second it remains held to the tissues of the narrow corollary tube by an " action comparable to that which results from the introduction of the moistened finger into a tube, or, better, into the mouth of a bottle. " The first can hold bees as well as Lepidoptera, and the second catches only the great sphinx moths whose beak is long and bulky. In both cases the insects often escape. The victims of the Asclepiads die of inanition without causing any damage to the flower. The large sphinx moths, however, thresh around before dying, and mutilate or tear off the flowers of the Hedychium.
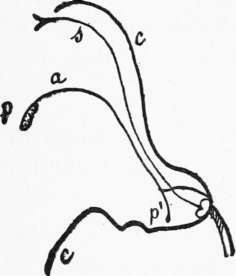
Fig. 7.-Vertical section of the flower of sage. In introducing itself into the corolla c to reach the nectaries situated at the bottom, the bee pushes aside the little arm p' from the connection with the stamen a and receives on its back the contents of the pollen sac p situated at the end of the large arm; when it afterwards penetrates a flower it scrapes with the back the bifid stigma of the style s which charges it with pollen.
Traps of another sort have been described by H. Miiller in the butterwort of the Alps, and by Sprengel in the Aristolochias. In these last plants the hairs which transform the long floral tube into a net do not stay rigid for a long time ; they wither at the moment when the pollen sacs in the chamber which terminates the tube open, and the insects captured in this chamber can then go out, carrying with them the grains of pollen which will fertilize other flowers.
From the fact that certain trap plants hold and kill some of the insects which visit them, we may conclude that the reciprocal adaptation is a lure. These plants show us simply that the best adaptation cannot fit every case; all flowers are not adapted to all insects, and reciprocally; but there are enough hospitable flowers to save the race of insects which may take themselves to the trap flowers, and enough insects to bring about without injury the fertilization of the latter. I add that it is natural that the victims of the trap plants are not adapted to play with the trap, for adaptation presupposes heredity and these insects will not leave any descendants.
In reciprocal adaptation each adapted one works for its own benefit without regard to the other, and if the plant has only indifference for the insect, the latter also is indifferent to the fate of the plant. Osmia pérezi decorates its nest with strips torn from the petals of Convolvulus cantabrica and C. althceoides, but the flowers torn by the insect are ruined and are destined to be sterile if they are not fertilized in advance. "I marked fifteen flowers torn by the bee," says Ferton, "and . . . six hours later I found certainly eight more, of which only one had pollen grains ; the other seven were dried, sterile, victims of their beauty.7 9
It is necessary to apply to reciprocal adaptation the aphorism of Claude Bernard, "The law of physiological finality is in each being, and not outside ; the living organism . . . works for itself and not for others."
Continue to: